Light is vital for photosynthesis, but is also necessary to direct plant growth and development. Light acts as a signal to initiate and regulate
photoperiodism and
photomorphogenesis. There are two light-sensing systems involved in these responses, the
blue light sensistive system and the red light sensitive or
phytochrome system.
Blue light responses: Many plant responses are regulated by blue light, including
phototropism, stomatal opnening and
chlorophyll synthesis. The last step of chlorophyll synthesis requires high levels of blue light. The other blue light responses are triggered by lower levels of blue light. For more detailed information, read [ this link ].
Phytochrome responses: Important plant responses regulated by the phytochrome system include photoperiodic
induction of flowering,
chloroplast development (not including chlorophyll synthesis), leaf
senescence and leaf
abscission.
Characteristics of phytochrome-mediated responses:
- The action spectrum of the light needed for these responses shows a peak in the red at about 660 nm.
- These responses can be reversed by an application of far-red light (peak at 730 nm) soon after the red treatment.
- Sensitive spectrophotometers can measure a decrease in absorbance at 660 nm and in increase in absorbance at 730 nm when sensitive plant tissues are exposed to red (660 nm) light.
- The change in absorbance is caused by the conversion of a the photoreceptor from one structural form to another. The red-absorbing form changes to the far-red absorbing form when it absorbs red light (660 nm) and back again when it absorb far-red light (730 nm).
The phytochrome molecule is the photoreceptor for red light responses. It exists in two forms, Pr and Pfr:
The Pr form:
- Absorbs at a peak of 666 nm
- Is the form synthesized in dark-grown seedlings.
- When Pr absorbs red light, it is converted to the Pfr form.
The Pfr form:
- Absorbs at a peak of 730 nm
- The Pfr form is the active form that initiates biological responses
- When Pfr absorbs far red light, it is converted to the Pr form
- Pfr can also spontaneously revert to the Pr form in the dark over time = dark reversion; Pfr is also susceptible to proteinases.
- Pfr absorbs some red light, so in red light, there is a balance of 85% Pfr and 15% Pr
- Pr absorbs very little far red light, so in far red light, there is a balance of 97% Pr to 3% Pfr
Phytochrome is a family of proteins with a small covalently-bound pigment molecule:
- Phytochrome proteins occur as a dimer of two identical 124 kDa polypeptides, each with a covalently-attached pigment molecule. Each polypeptide has three segments, or domains that have amino acid sequences similar to domains of other sensor proteins. [ reference ]
- The pigment is called the chromophore. It is a linear tetrapyrrole.
- When the chromophore absorbs light, there is a slight change in its structure. This causes a change in the conformation of the protein portion to the form that initiates a response.
- Phytochrome levels are much higher (about 50X) in dark-grown seedlings than in light-grown plants. Its levels are highest near the apex of the plant.
- Molecular genetics has revealed the existence of several genes for this protein in a given plant. All of these proteins use the same chromophore but differ in their sensitivity to light.
- The different phytochromes are involved in different biological responses to red light.
- Read more about the phytochrome molecule
Phytochrome allows plants to sense the color of light.
- Sunlight has a R:FR ratio of 1.2
- Light under a canopy of leaves has a R:FR ratio of 0.13
- Light under 5 mm of soil has a R:FR ratio of 0.88
- A higher proportion of FR light allows plants to detect when they are shaded.
- Plants adapted for growth in full sun will display greater stem elongation when they are transferred to shade. They also develop smaller leaves and less branching. This change is due to greater proportion of Pr to Pfr.
- Seeds of certain plants require red light for germination; FR light inhibits germination. Many small seeds with low amounts of storage reserves (such as lettuce) show such a red light requirement.
- If these seeds they are buried below the level of light penetration in the soil, they do not germinate.
- If they are shaded by a leaf canopy, causing a high proportion of FR, germination is inhibited, Pfr is required for germination.
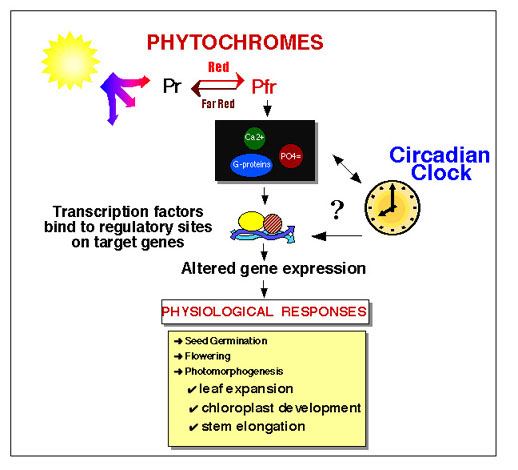
Phytochrome and the Circadian Clock in Plants
"
Photoreceptors and circadian clocks are universal mechanisms for sensing and responding to the light environment. In addition to regulating daily activities, photoreceptors and circadian clocks are also involved in the seasonal regulation of processes such as flowering. Circadian rhythms govern many plant processes, including movements of organs such as leaves and petals, stomata opening, stem elongation, sensitivity to light of floral induction, metabolic processes such as respiration and photosynthesis and expression of a large number of different genes." - drawing and quote from
Elaine Tobin's Website, UCLA.
reference ]
- Phytochrome A (PhyA), present only in angiosperms, is responsible for early events in germination and seedling de-etiolation. It is powerfully down-regulated in light both at the transcriptional and post-translational levels. In darkness it accumulates to (comparatively) high levels.
Expression of the other phytochrome types (B to E in angiosperms) is independent of light and both Pr and Pfr forms are stable.
- Phytochrome B (PhyB) is probably the photoreceptor involved in shade detection and avoidance. This response allows many species to greatly increase their stem extension rate when they become shaded by competitors. The relative amount of Pfr is reduced strongly by the presence of chlorophyll-bearing leaves that filter-out red light but not far-red. The absolute irradiance is irrelevant. Through this red/far-red sensitivity, phytochrome provides the plant with a degree of color perception. PhyB also is considered responsible for daylength detection in flowering and for tuberization in the potato, though the mechanisms are not understood.
- Phytochrome C (phyC) is a low-abundance member of the five-membered phytochrome family of photoreceptors in Arabidopsis. Experimental data indicate that phyC may have some physiological roles that are different to those of phyA and phyB in the control of seedling responses to light signals.
The Elusive Phytochrome "Receptor"
The mechanism by which the phytochrome (phy) photoreceptor family
transduces informational light signals to photoresponsive genes is still unclear, although progress has been made.
- Phytochrome-GFP fusion proteins migrate to the cell nucleus after they are activated by red light.
- In the case of PhyB, both photoactivation and nuclear translocation combined are necessary and sufficient for biological function. Conversely, neither artificial nuclear translocation of non-photoactivated phyB nor artificial retention of photoactivated phyB in the cytosol provides detectable biological activity.
Several candidates for a phytochrom receptor are being investigated. For example:
- PIF3, a phytochrome-interacting factor necessary for normal photoinduced signal transduction, is a novel basic helix-loop-helix protein.
Ni M, Tepperman JM, Quail PH. Cell 1998 Nov 25;95(5):657-67
The mechanism by which the phytochrome (phy) photoreceptor family transduces informational light signals to photoresponsive genes is unknown. Using a yeast two-hybrid screen, we have identified a hytochrome-interacting factor, PIF3, a basic helix-loop-helix protein containing a PAS domain. PIF3 binds to wild-type C-terminal domains of both phyA and phyB, but less strongly to signaling-defective, missense mutant-containing domains. Expression of sense or antisense PIF3 sequences in transgenic Arabidopsis perturbs photoresponsiveness in a manner indicating that PIF3 functions in both phyA and phyB signaling pathways in vivo. PIF3 localized to the nucleus in transient transfection experiments, indicating a potential role in controlling gene expression. Together, the data suggest that phytochrome signaling to photoregulated genes includes a direct pathway involving physical interaction between the photoreceptor and a transcriptional regulator.
- Phytochrome B protein binds to a clock protein, ADO1
"An Arabidopsis circadian clock component interacts with both CRY1 and phyB"
JOSE A. JARILLO, JUAN CAPEL, RU-HANG TANG, HONG-QUAN YANG, JOSE M. ALONSO, JOSEPH R. ECKER & ANTHONY R. CASHMORE. Nature 410, 487 - 490 (2001)
Most organisms, from cyanobacteria to mammals, use circadian clocks to coordinate their activities with the natural 24-h light/dark cycle. The clock proteins of Drosophila and mammals exhibit striking homology but do not show similarity with clock proteins found so far from either cyanobacteria or Neurospora. Each of these organisms uses a transcriptionally regulated negative feedback loop in which the messenger RNA levels of the clock components cycle over a 24-h period. Proteins containing PAS domains are invariably found in at least one component of the characterized eukaryotic clocks. Here we describe ADAGIO1 (ADO1), a gene of Arabidopsis thaliana that encodes a protein containing a PAS domain. We found that a loss-of-function ado1 mutant is altered in both gene expression and cotyledon movement in circadian rhythmicity. Under constant white or blue light, the ado1 mutant exhibits a longer period than that of wild-type Arabidopsis seedlings, whereas under red light cotyledon movement and stem elongation are arrhythmic. Both yeast two-hybrid and in vitro binding studies show that there is a physical interaction between ADO1 and the photoreceptors CRY1 and phyB. We propose that ADO1 is an important component of the Arabidopsis circadian system.